
Ceramides
Ceramides are lipids and they belong to the sphingolipid class. Ceramides are essential structural part of cell membrane (Maggio et al. 2006), however ceramides are also bioactive signaling lipids participating in many biologically crucial functions. Ceramides participate in regulation of many cellular and biochemical functions such as cell differentiation and proliferation, cell adhesion, inflammation, cell stress, and apoptosis (programmed cell death). (Sasset et al. 2016, Rodriquez-Cuenca et al. 2017, Borodzicz et al. 2015)
Ceramides are Carried by Lipoproteins
In circulation, ceramides are carried by different lipoproteins: VLDL, LDL, HDL and IDL. Ceramides are located on the surface of lipoprotein particles together with phospholipids, unesterified cholesterol, and apolipoproteins. (Kontush et al. 2013)
Ceramides are synthesized in all tissues, but the main ceramide source is liver, where ceramides are packed into lipoproteins. Ceramides are synthesized also in peripheral organs, from where ceramides are transported into circulation by small-sized HDL-lipoprotein.
Concentration of ceramides between different lipoproteins varies. Usually most of the ceramides are located in LDL, one fourth in HDL and one sixth in VLDL. (Wiesner et al. 2009, Ståhlman et al. 2013)
Ceramides and Atherosclerosis
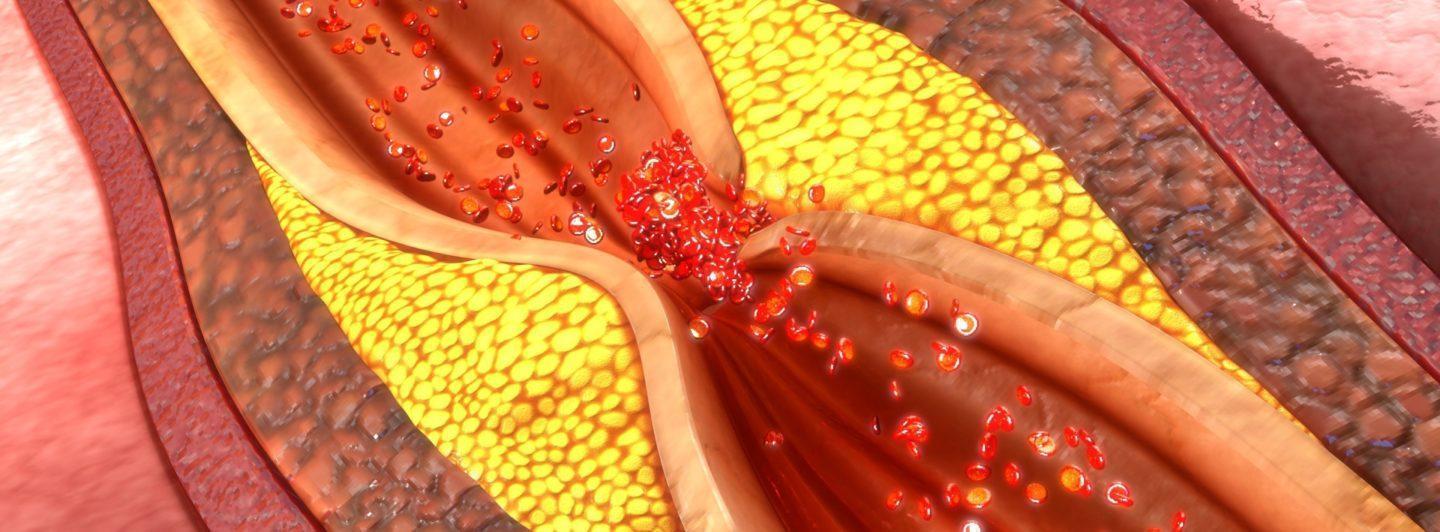
Ceramides have significant role in the development of unstable atherosclerotic cardiovascular disease. Ceramides accumulate in the atherosclerotic plaques and are involved in the formation of unstable plaques. (Schissel et al. 1996, Li et al. 2014)
Although the role of ceramides in cardiovascular diseases have been investigated lately, there is still much that is not known on their specific functions. It has been shown that ceramides are involved in many atherosclerotic processes such as lipoprotein aggregation, lipoprotein and cholesterol infiltration into macrophages and into the vessel wall (Devlin et al. 2008, Li et al. 2014, Schissel et al. 1996), mitochondrial dysfunction, platelet activation, and regulation of cytokines (Chaurasia & Summers 2015).
Furthermore, ceramides regulate signaling pathways which control blood pressure and liver steatosis, thus, being part of processes preceding cardiovascular events (Chaurasia & Summers 2015). Some of the ceramides may activate cardiomyocyte specific processes causing lipotoxic cardiomyopathy (Park & Goldberg 2012).
Inflammatory cytokines such as gamma-interferon, TNF-alfa, and interleukin-1 beta, stimulate ceramide synthesis (Marathe et al. 1998) by activating ceramide synthesizing enzymes sphingomyelinases (Bikman 2012, Bikman & Summers 2012, Holland et al. 2011). These ceramide syntheses activating cytokines participate in inflammatory processes, which are instrumental part in the development process of atherosclerotic events (Verma et al. 2017).
Ceramides in Atherosclerotic Plaques
In circulation, ceramides participate in the process of lipoprotein and cholesterol aggregation and infiltration into the vessel wall thus advancing the plaque formation (Li et al. 2014). Ceramides also activate platelets and induce endothelial dysfunction in the plaque development (Knapp et al. 2012, Zhang et al. 2012, Mugabo et al. 2011).
Ceramides are enriched in arterial plaque by as much as 50-fold (Schissel et al. 1996, Edsfeldt et al. 2016). This is likely due to the inflammatory cytokines in the plaques. Plaque ceramides in turn activate local inflammatory pathways thus advancing the development of atherosclerosis even more (Bismuth et al. 2008, Edsfeldt et al. 2016). It has been shown by imaging technologies that ceramides are especially found from atherosclerotic plaques with necrotic core (Uchida et al. 2017).

Literature on Ceramides
Maggio et al. Biophysics of sphingolipids II. Glycosphingolipids: an assortment of multiple structural information transducers at the membrane surface. Biochim Biophys Acta 2006; 1758: 1922-44.
Sasset et al. Sphingolipid de novo biosynthesis: a rheostat of cardiovascular homeostasis. Trens Endocrinol. Metab. 2016; 27: 807-19.
Rodriguez-Cuenca et al. Sphingolipids and glycerophospholipids – The “ying and yang” of lipotoxicity in metabolic diseases. Prog. Lipid Res. 2017; 66:14-29.
Borodzicz et al. 2015 Sphingolipids in cardiovascular diseases and metabolic disorders. Lipids Health Dis. 2015; 14:55.
Kontush et al. Unraveling the complexities of the HDL lipidome. J Lipid Res. 2013; 54: 2650-63.
Wiesner et al. Lipid profiling of FPLC-separated lipoprotein fractions by electrospray ionization tandem mass spectrometry. J Lipid Res. 2009; 50:574-85.
Ståhlman et al. Dyslipidemia, but not hyperglycemia and insulin resistance, is associated with marketed alterations in the HDL lipidome in type 2 2 diabetic subjects in the DIWA cohort: impact on small HDL particles. Biochim Biophys Acta 2013; 1831: 1609-17.
Schissel et al. Rabbit aorta and human atherosclerotic lesions hydrolyze the sphingomyelin of retained low-density lipoprotein. Proposed role for arterial-wall sphingomyelinase in subendothelial retention and aggregation of atherogenic lipoprotein. J Clin. Invest. 1996; 98:1455-1464.
Li et al. Endogenous ceramide contributes to the transcytosis of oxLDL across endothelial cells and promotes its subendothelial retention in vascular wall. Oxid. Med. Cell Longev. 2014; 2014:823071.
Chavez & Summers A ceramide-centric view of insulin resistance. Cell Metab, 2012, 15(5):585-94
Markgraf et al. Lipidomics – Reshaping the analysis and perception of type 2 diabetes. Int. J. Mol. Sci. 2016 17:E1841.
Park & Goldberg Sphingolipids, lipotoxic cardiomyopathy, and cardiac failure. Heart Fail. Clin. 2012; 8: 633-641.
Chaurasia & Summers Ceramides – lipotoxic inducers of metabolic disorders. Trends Endocrinol. Metab. 2015; 26: 538-550.
Devlin et al. Acid sphingomyelinase promotes lipoprotein retention within early atheromata and accelerates lesion progression. Arterioscler. Thromb. Vasc. Biol. 2008; 28: 1723-1730.
Bikman. A role for sphingolipids in the pathophysiology of obesity-induced inflammation. Cell Mol. Life Sci. 2012; 69:2135-2146.
Bikman & Summers. Ceramides as modulators of cellular and whole-body metabolism. J. Clin. Invest. 2011; 121:4222-4230.
Holland et al. Receptor-mediated activation of ceramidase activity initiates the pleiotropic actions of adiponectin. Nat. Med. 2011; 17: 55-63.
Verma et al. CANTOS ushers in a new calculus of inflammasome targeting for vascular protection – and maybe more. Cell Metab. 2017; 26: 703-705.
Bismuth et al. 2008. Ceramide: A Common Pathway for Atherosclerosis? Atherosclerosis. 2008; 196(2): 497–504
Edsfeldt et al. Sphingolipids contribute to human atherosclerotic plaque inflammation. ATVB 2016; 36:1132-40. Uchida et al. Detection of Ceramide, a Risk Factor for Coronary Artery Disease in Human Coronary Plaques by Fluorescent Angioscopy. Circ. J. 2017; 81: 1886-1893.
Knapp et al. Myocardial infarction differentially alters sphingolipid levels in plasma, erythrocytes and platelets of the rat. Basic Res. Cardiol. 2012; 107: 294.
Zhang et al. Ceramide mediates vascular dysfunction in diet-induced obesity by pp2a-mediated dephosphorylation of the enos-akt complex. Diabetes 2012; 61:1848-1859.
Mugabo et al. Palmitate induces C-reactive protein expression in human aortic endothelial cells. Relevance to fatty-acid induced endothelial dysfunction. Metabolism. 2011; 60:640-648.